
При всей важности абиотических условий определяющее значение во взаимодействии биоты и среды обитания имеют биотические факторы с доминантной ролью трофического. Трофические взаимосвязи в сообществах различных участков Байкала исследованы недостаточно полно. К настоящему времени наиболее детально изученной является только пелагиаль озера, характеризующаяся относительно невысоким видовым разнообразием доминирующих на различных трофических уровнях организмов и высокой численностью и биомассой отдельных видов. Ихтиомасса пелагических рыб составляет более 80% от общей ихтиомассы рыб озера. Биомасса желтокрылки в современный период определяется в 10–12 тыс. т, длиннокрылой широколобки в 3 тыс. т, малой голомянки от 61,5 до 103 тыс. т, большой голомянки от 73,3 до 112 тыс. т, а омуля от 30 до 40 тыс. т.
Основной поток энергии в экосистеме Байкала идет через планктонную сеть пелагиали, в которой трансформируется до 99% энергии. Продукция различных трофических уровней в этой зоне представлена в таблице 2.10.
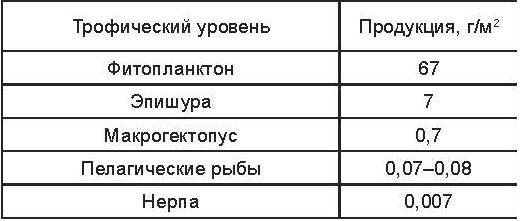
Таблица 2.10. Величины продукции (г/м2) разных трофических уровней в пелагиали Байкала.
Прибрежно-соровая зона. Схема потока вещества и энергии прибрежно-соровой зоны представлена на рис. 2.66. Внутренние мелководные участки крупных заливов, водоемы дельты р. Селенги, нижнее течение крупных рек и озера, расположенные в участках прибрежно-соровой зоны, населяют щука, плотва, елец, язь, сазан, карась, озерный гольян, окунь, песчаная широколобка. Молодь этих видов на первом году жизни обитает на хорошо прогреваемых и зарастающих высшей водной растительностью мелководьях. В течение первого года жизни сеголетки питаются преимущественно организмами зоопланктона. Только щука начинает хищничать уже в месячном возрасте, поедая личинок других видов рыб. Соответственно, наибольшая напряженность пищевых взаимоотношений у молоди этих видов возникает при питании зоопланктоном. В старшем возрасте основу питания рыб прибрежно-сорового комплекса составляют амфиподы, моллюски и личинки амфибиотических насекомых (все виды рыб), водная растительность (плотва, язь, сазан) и рыба (щука, окунь). Наиболее высокая вероятность конкурентных отношений наблюдается между плотвой, язем и сазаном при питании водной растительностью, моллюсками и личинками ручейников. Экспансия и дальнейшая натурализация ротана в дельте Селенги и прилежащих районах в значительной мере нарушила характер исторически сложившихся здесь пищевых взаимоотношений. Установлено, что наибольшая напряженность пищевых отношений возникает между ротаном и язем (сходство состава пищи достигает 90%), ротаном и карасем (81,2%), ротаном и плотвой (67,3%). Высокая численность ротана в дельте р. Селенги является мощным прессом, снижающим численность частиковых видов рыб. Это, в свою очередь, привело к значительным изменениям в питании хищных видов рыб и рыбоядных птиц, практически полностью перешедших на питание ротаном. Наличие многочисленной и легко доступной жертвы увеличило темп роста и плодовитости щуки и окуня, численность ротана в последние годы снижена.
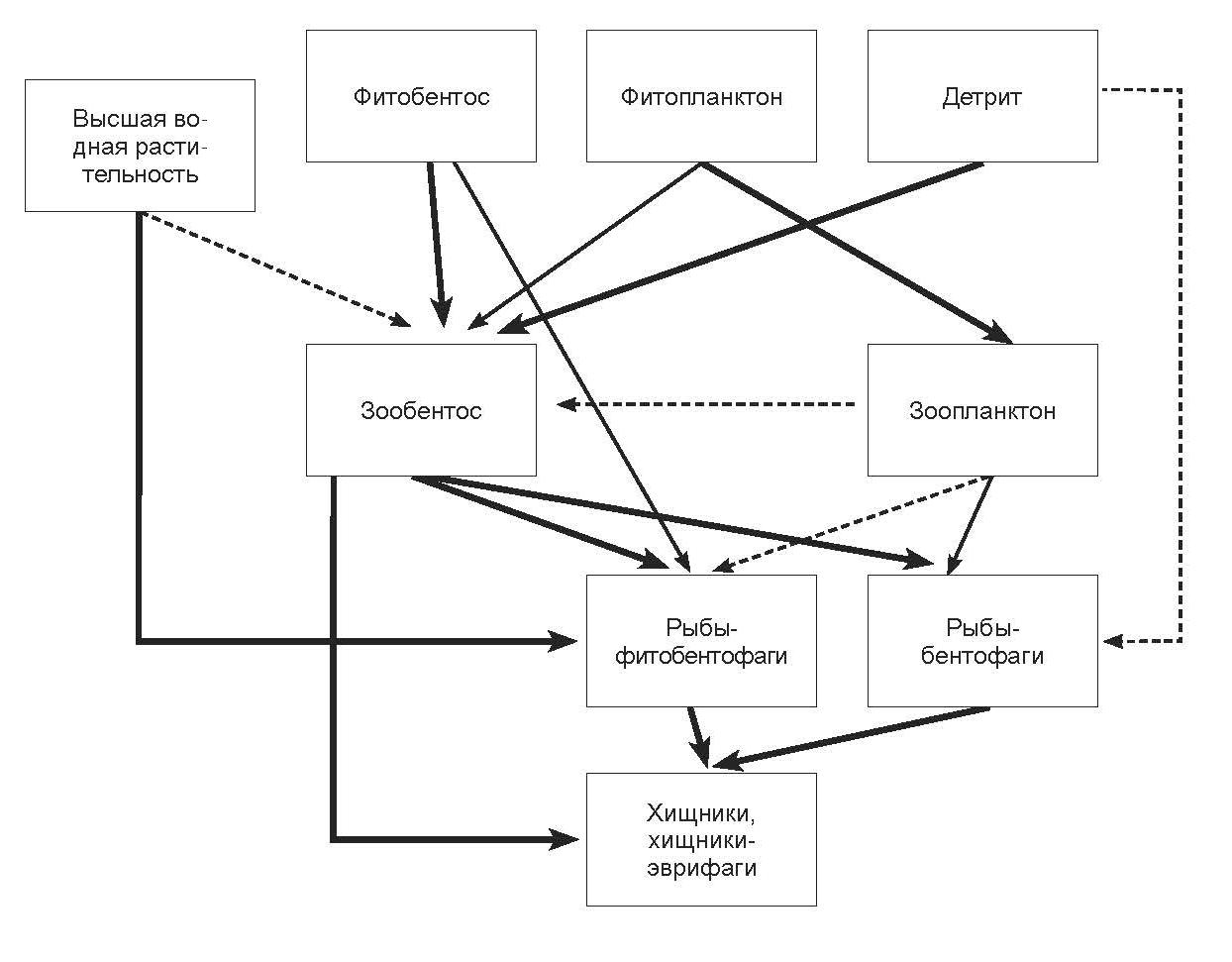
Рис. 2.66. Упрощенная схема потока энергии в прибрежно-соровой зоне озера Байкал.
Литораль. Схема потока вещества и энергии в зоне литорали представлена на рис. 2.65. В литорали озера и предустьевых участках небольших горных притоков озера в зоне глубин от уреза воды до 20–30 м и несколько глубже основными группами организмов, используемыми в пищу рыбами, являются амфиподы (все виды рыб), моллюски (озерноречной сиг-пыжьян, байкальский озерный сиг), ручейники (черный байкальский хариус, белый байкальский хариус, озерно-речной сиг-пыжьян, байкальский озерный сиг, ленок), хирономиды (озерно-речной сиг-пыжьян, байкальский озерный сиг, черный байкальский хариус, белый байкальский хариус, песчаная широколобка), воздушно-наземные насекомые (черный байкальский хариус, белый байкальский хариус) и рыба (таймень, ленок, черный байкальский хариус, белый байкальский хариус, налим, байкальская большеголовая широколобка). Анализ спектров питания рыб, населяющих литораль озера, показал, что в их рационе преобладают амфиподы из разных систематических и экологических групп. При этом состав этих групп специфичен у каждого вида рыб.
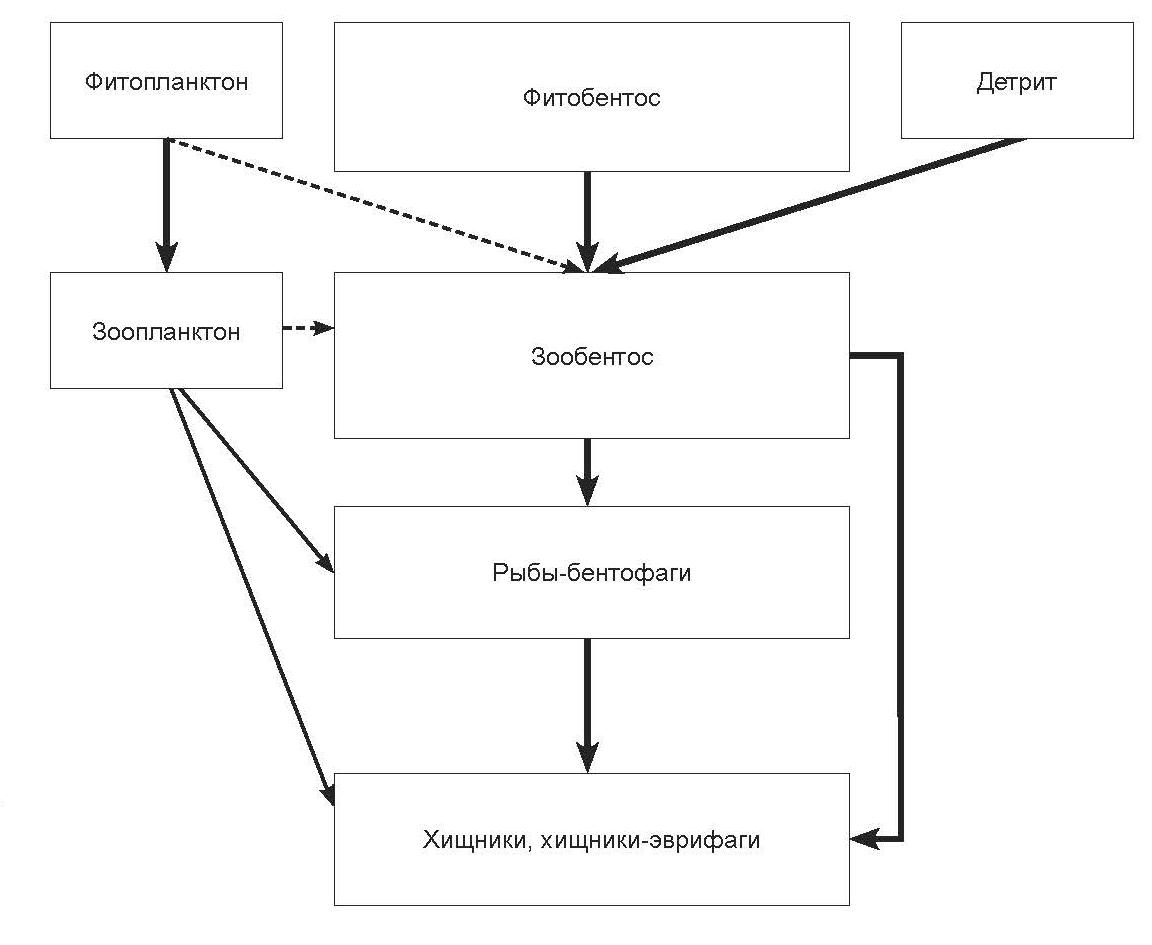
Рис. 2.67. Упрощенная схема потока энергии в литорали озера Байкал.
Общими являются преимущественно второстепенные компоненты, что обусловливает низкую степень конкуренции за пищевые ресурсы. Сходство состава пищи между видами рыб, населяющих литораль Байкала, не превышает 20%.
Абиссаль. Схема потока вещества и энергии в зоне абиссали представлена на рис. 2.68. Эндемичные рогатковидные рыбы по пищевой специализации являются бентофагами и хищникамиэврифагами. Ранее существовало мнение о том, что эволюция рогатковидных рыб в глубинных водах Байкала шла путями эволюции их жертв – невооруженных, или «гладких», бокоплавов, обусловленной расселением последних по разным биотопам и экологическим нишам.

Рис. 2.68. Упрощенная схема потока энергии в абиссали озера Байкал.
Исследованиями последних лет показано, что помимо рыб, питающихся невооруженными амфиподами, имеется ряд видов (пестрокрылая, жирная, широкорылая, плоская, узкая, большая широколобки), которые во взрослом состоянии переходят на преимущественное потребление крупных вооруженных видов бокоплавов, ведущих как донный, так и батипелагический образ жизни. В питании этих же видов в значительном количестве также отмечаются рыбные жертвы, представленные преимущественно видами пелагического комплекса – малой и большой голомянками, желтокрылкой и длиннокрылкой.
Разнообразны у хищных видов рогатковидных рыб и способы охоты за своими жертвами. Жирная широколобка, имеющая нейтральную плавучесть и парящая в придонных слоях воды, захватывает проплывающих в непосредственной близости голомянок и амфипод. Пестрокрылая, широкорылая и большая широколобки ведут малоподвижный образ жизни и подстерегают свои жертвы на дне водоема. Плоская и узкая широколобки являются так называемыми хищниками-рейдерами, активно передвигающимися в придонных слоях воды и разыскивающими своих жертв. Снижение напряженности конкурентных взаимоотношений между видами рогатковидных рыб в глубоководной зоне осуществляется за счет их биотопической разобщенности, различий в способах добывания пищи и уникальных сочетаний видового состава потребляемых амфипод. Наблюдения, проведенные с подводных управляемых аппаратов «Pisces», свидетельствуют о низкой плотности рогатковидных рыб в абиссали и их слабой двигательной активности, что может служить дополнительным подтверждением мнения об отсутствии напряженности в пищевых взаимоотношениях рыб в абиссальной зоне Байкала.
Пелагиаль. Схема потока вещества и энергии в зоне пелагиали представлена на рис. 2.69. Исследования межгодовых изменений динамики качественных и количественных показателей зоопланктона в пелагиали выявили короткои долгопериодичную цикличность изменений структуры его сообщества в зависимости от особенностей развития фитопланктона. В настоящее время это обобщенно выражается в наличии трех качественно различных состояний планктона озера и соответственно названных типов лет – «предмелозирные», «мелозирные» и «послемелозирные» годы, характеризующиеся специфичным набором доминантных видов и особенностями сезонной сукцессии планктона. «Мелозирные» (аулякозейрные) годы характеризуются доминированием водорослей рода Aulacoseira (ранее Melosira) и депрессией численности эпишуры. C. kolensis в этот период имеет наиболее благоприятные условия для своего развития и доминирует в структуре зоопланктона пелагиали, особенно сильно это проявляется в теплые годы. «Послемелозирные» годы отличаются интенсивным развитием водорослей рода Synedra и летнее-осенней генерации эпишуры. В предмелозирные годы наибольшее развитие в планктоне имеют Cyclotella minuta и зимнее-весенняя генерация эпишуры. К наиболее бедным по эпишуре «мелозирным» годам относятся 1964, 1982, 1984, 1990 гг. Объектами питания эпишуры, помимо водорослей, являются бактерии и инфузории. Питание C. kolensis, начиная с третьей копеподитной стадии, основывается преимущественно на крупных животных объектах – эпишуре, коловратках, инфузориях, хотя могут использоваться и растительные. Установлено, что циклопы в годы их обилия играют значительную роль в регуляции численности эпишуры.
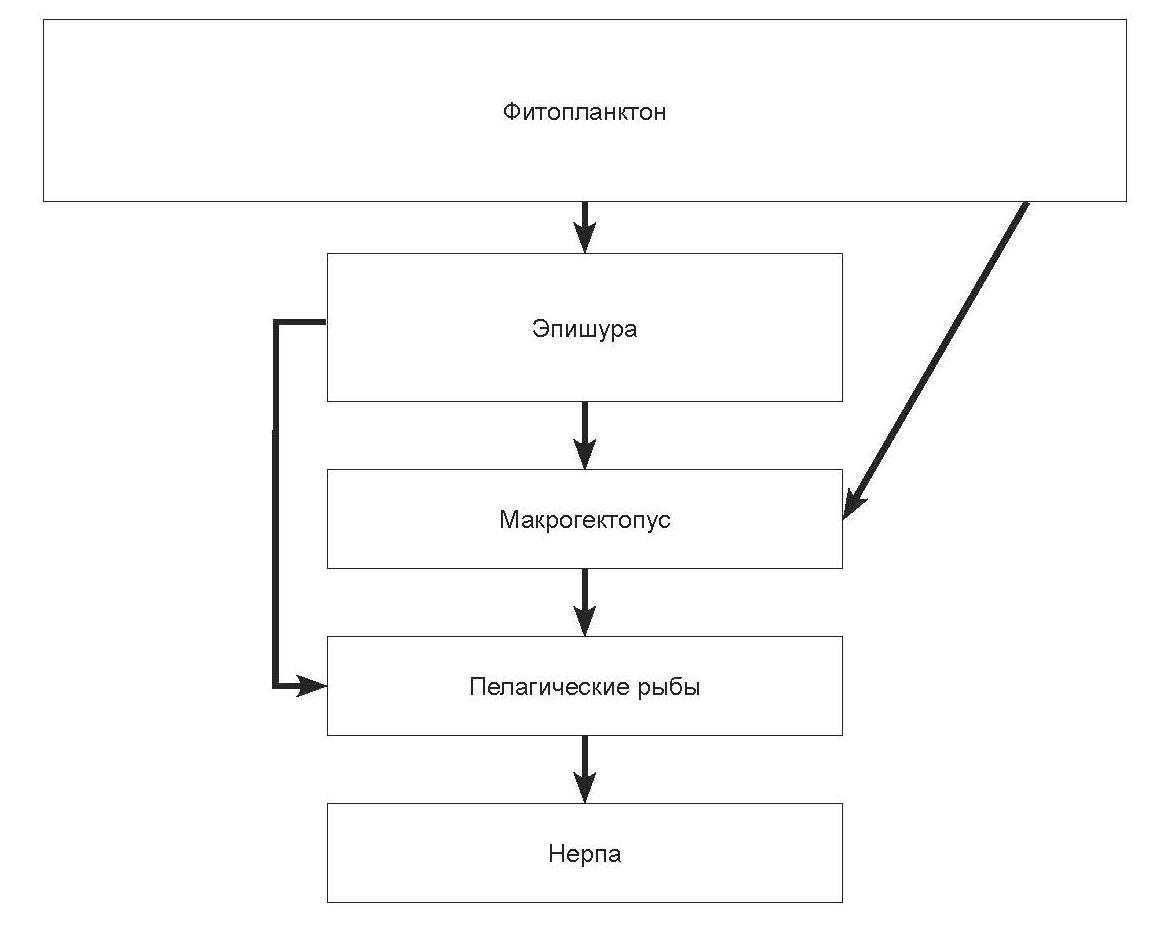
Рис. 2.69. Упрощенная схема потока энергии в пелагиали озера Байкал.
Значительное развитие ветвистоусых ракообразных отмечается не ежегодно и носит сезонный характер. Наиболее характерными видами среди этой группы в планктоне озера являются Daphnia galeata и Bosmina longirostris. В планктоне озера кладоцеры обычно появляются в конце июля–августе, а пик их развития приходится на сентябрь–октябрь, когда численность босмин достигает 133 тыс. экз./м3, дафний – 90 тыс. экз./м3. Значительные концентрации кладоцер отмечаются в осенний период в пелагиали прибрежий в районе крупных заливов, мелководий и бухт.
Важную роль в сообществе пелагиали и ее пищевых цепях играет батипелагический бокоплав макрогектопус, питающийся фитои зоопланктоном. Являясь первичным и вторичным консументом, он в значительной мере аккумулирует органическое вещество предыдущих трофических уровней, переносит его преимущественно к шести видам пелагических рыб и в гораздо меньшем количестве – опускаясь в придонные слои к абиссальным рогатковидным рыбам.
Макрогектопус обитает, в основном, в глубоководной части озера, в районах с глубинами менее 100 м он встречается эпизодически. У макрогектопуса четко выражены суточные вертикальные миграции: в ночное время отмечается подъем бокоплавов в верхние богатые планктоном слои воды. Особенностью пространственного распределения макрогектопуса (юра) в озере является приуроченность наибольших скоплений его к крутым склонам в районах, примыкающих к крупным заливам и обширным мелководьям. К таковым можно отнести внешние части Баргузинского и Чивыркуйского заливов, северный створ Малого Моря, периферийную часть Селенгинского мелководья, а также внешнюю часть острова Ольхон. Максимальная численность макрогектопуса в районе Среднего Байкала у мыса Хобой достигала 6250 экз./ м2 над максимальными глубинами. В Южном и Северном Байкале отмечаются в 2–3 раза меньшие значения его численности. Гидроакустическими исследованиями установлена двуслойность распределения вида. Его скопления в верхних слоях (50–150 м) сформированы мелкоразмерными особями обоих полов (5–6 мм), а в нижних слоях (100–200 м) они преимущественно сформированы крупными самками. Максимальные концентрации в этих скоплениях, по данным гидроакустических исследований, могут достигать 6–8 г/м3.
По мнению ряда исследователей, основное влияние на численность эпишуры и, вероятно, циклопа в литорали озера оказывает выедание их хищными беспозвоночными и молодью рыб, значительно меньше влияют взрослые рыбы-планктофаги.
Существуют две противоположные точки зрения по поводу использования зоопланктона пелагическими рыбами и кормовой емкости пелагиали. Согласно первой, зоопланктон недоиспользуется пелагическими рыбами при их современной численности, и в связи с этим возможна акклиматизация в оз. Байкал ряда других планктоноядных рыб. Согласно второй, которой в настоящее время придерживается большинство исследователей, в пелагиали уже с первого трофического уровня возникает значительная напряженность в пищевых отношениях, и не существует целесообразности в перестройке ихтиоценоза пелагиали озера.
Основными кормовыми организмами для омуля являются эпишура, макрогектопус, молодь пелагических рогатковидных рыб и донные амфиподы, для большой голомянки – макрогектопус и малая голомянка, для малой голомянки – эпишура и макрогектопус, для длиннокрылки – макрогектопус, молодь пелагических рогатковидных рыб и донные амфиподы, для желтокрылки – эпишура, циклопы и дафнии, а также частично молодь донных амфипод. Высокая степень сходства состава пищи, обусловленная малым видовым разнообразием кормовых объектов в пелагиали, наряду с высокой численностью рыб пелагического комплекса служат предпосылками возникновения напряженности в пищевых отношениях между ними. Наибольшее пищевое сходство при питании эпишурой отмечается между омулем и желтокрылкой в прибрежных участках пелагиали и малой голомянкой и омулем в пелагиали открытой части озера. Установлено, что омуль переходит на питание эпишурой лишь при значительных концентрациях последней, тогда как желтокрылка питается эпишурой при ее низких концентрациях и частично переходит на питание другими объектами при повышении ее численности, тем самым избегая конкуренции с омулем. При питании макрогектопусом сходство состава пищи отмечается между омулем и длиннокрылкой, омулем и голомянками, длиннокрылкой и голомянками. Напряженность в пищевых взаимоотношениях смягчается приспособлением отдельных видов к различным оптимальным для питания температурам (для голомянок это температуры ниже 8оС, для длиннокрылки и северобайкальской желтокрылки – 10–12оС, для желтокрылки – 12–15о С) и связанным с этим распределением по разным глубинным горизонтам, а также особенностями распределения основных объектов их питания и переходом на питание ихтиопланктоном омуля, длиннокрылки и большой голомянки.
Замыкает трофическую пирамиду пелагиали оз. Байкал байкальская нерпа. При довольно широком общем спектре питания (около 30 видов рыб) абсолютным доминантным видом его во все периоды годового физиологического цикла является малая голомянка (от 40 до 90% по массе). Второе место принадлежит субдоминанту – длиннокрылке (от 4 до 39%). Большая голомянка (5–7%) и желтокрылка (0,5–10%) являются вторичными компонентами в рационе нерпы. Таким образом, нерпа утилизирует биологическую продукцию пелагиали Байкала. Литоральные виды широколобок (песчаная, каменная и большеголовая) составляют минимальную часть рациона в летне-осенний период.
В общем круговороте вещества и энергии в экологической системе Байкала до сих пор не дана оценка роли амфипод-трупоедов (стервятников), особенно рода Ommatogammarus. Несомненно, что роль его в деструкции органического вещества весьма и весьма значительная. Не случайно в Байкале практически не встречаются трупы погибших животных, даже через несколько дней после гибели, в результате аэромоноза окуня и щуки. Исключением из этого правила является только вынос на берег погибших тюленей при штормовой погоде.
Источник: Байкал: природа и люди : энциклопедический справочник / Байкальский институт природопользования СО РАН ; [отв. ред. чл.-корр. А. К. Тулохонов] – Улан-Удэ : ЭКОС : Издательство БНЦ СО РАН, 2009. С. 178-183.
Энциклопедии городов | Энциклопедии районов | Эти дни в истории | Все карты | Всё видео | Авторы Иркипедии | Источники Иркипедии | Материалы по датам создания | Кто, где и когда родился | Кто, где, и когда умер (похоронен) | Жизнь и деятельность связана с этими местами | Кто и где учился | Представители профессий | Кто какими наградами, титулами и званиями обладает | Кто и где работал | Кто и чем руководил | Представители отдельных категорий людей
